Trilobite Evolution
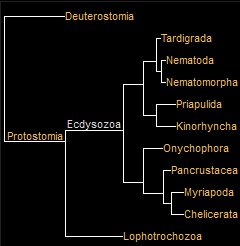
Trilobite OriginsThe story of trilobite evolution must begin with that of the arthropods. Trilobites were a member of what is far and away the largest phylum of animals, Arthropoda, with well more than a million described species of out of a total of perhaps 5 to 10 million extant, comprising around 80% of all animal species (Ødegaard, 2000). Prior to modern sequencing, scientists can only conjecture as to the characteristics of the first common ancestor of arthropods, but it surely lived in the Precambrian, a period for which the fossil record is exceedingly sparse and enigmatic. The evolutionary origins of trilobites is unsettled science, with the distinct possibility it always will be. Consensus exists that their origins were in the Precambrian time among the diverse bilaterians, probably in the Ediacaran Period (Lieberman, 2002). Their ancestors (or a common ancestor are likely among from the early Ediacaran arthropods, that are often labeled arachnomorphs or trilobitamorphs in the early Cambrian. A place to begin is the ancestory and sequencing-based phylogeny of Ecdysozoa, the moulting animals, among which is trilobita. Ecdysis unites trilobites with arthropodsFrom
phylogenetic analyses constructed using 18S
ribosomal RNA genes, Aguinaldo et
al., 1997 defined Superphylum Ecdysozoa
as a monophyletic clade
uniting eight phyla, arthropoda, tardigrades
and
onychophorans
that
Through modern sequencing, there is strong support combining the phyla Arthropoda, Tardigrada, and Onychophora into the unranked taxon Panarthropoda within superphylum Ecdysozoa. The phyla are united by common characteristics of ventral nervous system, a segmented body plan, and legs and claws. Ediacaran animals Parvancorina and Spriggina, García-Bellido, D. C.; Collins, D. H. (2004) Marrella from Burgess Shale earliest unequivocal evidence of moulting. [ (May 2004), Moulting arthropod caught in the act", Nature 429 (6987): 4] Superphylum: Ecdysozoa |
||||||||||||||||||||||||
Examples of Trilobite Evolution |
||||||||||||||||||||||||
The old saying goes, "use it or lose it". Charles Darwin spoke about seconadary blindness (progressive loss of vision through natural selection) in Origin of Species:
|
||||||||||||||||||||||||
| Size matters. Size matters a lot in evolution. But, it’s the species that can best adapt to its environment, particularly a changing environment that survives. In survival, the benefits accrued by larger size do not always endow the animal with a survival advantage – size matters in both providing advantages and disadvantages – there are tradeoffs. Famed American paleontologist Edward Drinker Cope (Hone, et al., 2005) postulates (in what some call Cope’s rule) that population members tend to increase in body size over evolutionary time scales. While larger body size provides higher fitness for many reasons, there are also many disadvantages for both individual organisms and entire populations. Moreover, the fossil record suggests that clades of larger animals are more susceptible to extinction as generational time period is longer and fecundity is reduced, clearly the worst outcome for the bullies and brutes. Here are some pros and cons to size to consider:
Evolutionary fitness disadvantages:
It’s difficult to know which of the above affected trilobite size changes, and which were more important and when, but trilobites clearly exhibited changes in size over geologic time. Trilobites have a more than 700 fold adult size range, enormous by any standard. The minimum is on the order of one mm, and the maximum more than 700 mm. The largest trilobite found in the fossil record is Isotelus rex, an Ordovician Asaphid trilobite discovered in 1998 (Rudkin, 2003) near the Hudson Bay in Canada. Bell, et al (2012) studied trilobite size variation over geologic time (click for larger image):
That is, while trilobites exhibit a fast size increase from the lower Cambrian to middle Cambrian, as well as into the Ordovician, size decreases thereafter. There is a gradual size decline through the Silurian. An initial size increase in the early Devonian is attributed to evolution of spiny exoskeletons in some species (Chatterton, 2006), noting also that the Frasnian/Famennian mass extinction removed all predatory species among orders Phacopida and Lichida, leaving generally more diminutive trilobites through the Carboniferous and into the Permian, when trilobites all went extinct.
|
||||||||||||||||||||||||
 The
trilobite fossil record is among the best of all the animal
groups. It is not surprising then that it
sometimes reveals sequences of evolutionary transition, recording
the adaptation, or descent with modification, to new selective
pressures as a matter
of survival,
or perhaps record failure to adapt,
leading to extinction.
One great
example is the radiation of Baltic asaphid trilobites of family
Asaphidae from the region that is now Saint Petersburg,
Russia from late in the
lower
Ordovician and into the middle Ordovician. A hundred years
of collecting of these
strata has provided tremendous data set for The
trilobite fossil record is among the best of all the animal
groups. It is not surprising then that it
sometimes reveals sequences of evolutionary transition, recording
the adaptation, or descent with modification, to new selective
pressures as a matter
of survival,
or perhaps record failure to adapt,
leading to extinction.
One great
example is the radiation of Baltic asaphid trilobites of family
Asaphidae from the region that is now Saint Petersburg,
Russia from late in the
lower
Ordovician and into the middle Ordovician. A hundred years
of collecting of these
strata has provided tremendous data set for  cladistics analysis.
The region that has long yielded a tremendous diversity of
trilobites is
believed to have been an inland sea during the earlier Ordovician
that was cut off from the ocean to the west. At some point toward
the late Ordovician the inland sea again became connected to
the ocean. The resulting flow of sediment (turbidity) clouded
the water
and settled on the seafloor. Through some 50 foot of limestone
encompassing some two million years, some
Asaphidae lines of descent developed stalked eyes of ever increasing height, ostensibly
to
enable them to better spot either prey or predators, or both.
The figure shows selected species and lines of descent. Not
all lines
produced elevated eyes. The figure below depicts the two best
known: 1) Asaphus lepidurus to Asaphus expansus, which branches
into
Asaphus cornutus with moderate stalked eyes, and then to Asaphus
kowalewskii,
with very high eye stalks that almost seem unnatural. The other
branch from Asaphus expanses leads to three species with successively
higher eyes, Asaphus kotlukovi, Asaphus
punctatus and Asaphus
intermedius.
It also warrants mention that the period beginning in the early Ordovician
is at times called "The
Great Ordovician Radiation", or even the
"Ordovician Explosion", denoting an enormous diversification of marine
animals, that had been decimated in the Cambrian-Ordovician
Extinction Event. cladistics analysis.
The region that has long yielded a tremendous diversity of
trilobites is
believed to have been an inland sea during the earlier Ordovician
that was cut off from the ocean to the west. At some point toward
the late Ordovician the inland sea again became connected to
the ocean. The resulting flow of sediment (turbidity) clouded
the water
and settled on the seafloor. Through some 50 foot of limestone
encompassing some two million years, some
Asaphidae lines of descent developed stalked eyes of ever increasing height, ostensibly
to
enable them to better spot either prey or predators, or both.
The figure shows selected species and lines of descent. Not
all lines
produced elevated eyes. The figure below depicts the two best
known: 1) Asaphus lepidurus to Asaphus expansus, which branches
into
Asaphus cornutus with moderate stalked eyes, and then to Asaphus
kowalewskii,
with very high eye stalks that almost seem unnatural. The other
branch from Asaphus expanses leads to three species with successively
higher eyes, Asaphus kotlukovi, Asaphus
punctatus and Asaphus
intermedius.
It also warrants mention that the period beginning in the early Ordovician
is at times called "The
Great Ordovician Radiation", or even the
"Ordovician Explosion", denoting an enormous diversification of marine
animals, that had been decimated in the Cambrian-Ordovician
Extinction Event. |
||||||||||||||||||||||||
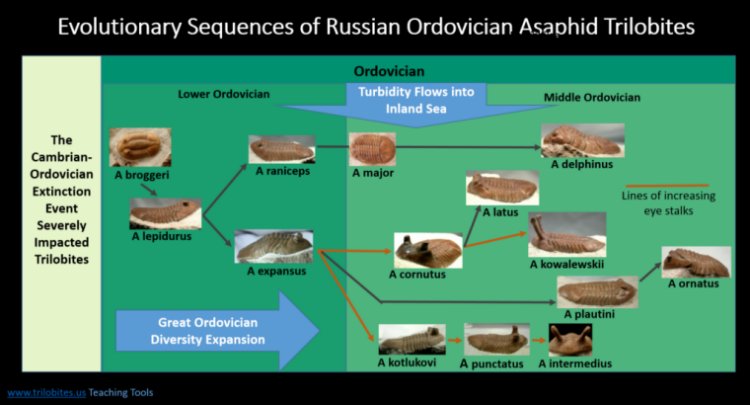 Figure above: Turbidity flows are selective pressure for trilobite in family Asaphidae to evolves eyes on ever higher stalks. Not all species in evolutionary lines are shown - click image to open new page with larger image. (Reference, Adamek, 2014) |
||||||||||||||||||||||||